|
Proszę chwilę zaczekać, ładuję stronę ... |
|
Proszę chwilę zaczekać, ładuję stronę ... |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
4.10. Metale ciężkie i siarka w porostach
W 2012 roku na 24 stanowiskach badawczych leżących w granicach zlewni eksperymentalnej (12 powierzchni) i w jej otulinie (12 pow.) pobrano próby plech porostu Hypogymnia physodes (L.) Nyl. Próby po wysuszeniu i opisaniu przesłane zostały do Zakładu Monitoringu Środowiska Instytutu Nauk o Środowisku Uniwersytetu Jagiellońskiego w Krakowie, gdzie poddane zostały badaniom na zawartość metali ciężkich – kadmu, ołowiu, miedzi, cynku, chromu, niklu i żelaza oraz siarki. Wyniki analiz chemicznych przedstawiono w tabeli 60, a ich opracowanie stanowi odrębną część Raportu, wykonaną przez Uniwersytet Jagielloński.
Tabela 60. Koncentracja (µg/g s.m.) Cd, Pb, Cu, Zn, Fe, Cr, Ni i S w plechach porostu Hypogymnia phsodes
4.11. Fauna epigeiczna
Badania dotyczące fauny epigeicznej dotyczyły tylko chrząszczy z rodziny biegaczowatych Carabidae i prowadzone były w dwóch typach siedliskowych lasu - w borze bagiennym i lesie mieszanym. Na każdym siedlisku wytypowano po trzy stanowiska oddalone od siebie o około 250 m, na których zainstalowano po 5 pułapek rozmieszczonych liniowo co trzy metry. Pułapki (słoiki o pojemności 0,33 l i średnicy otworu 56 mm) zostały wkopane równo z powierzchnią gruntu i osłonięte daszkami. Jako płynu konserwującego użyto roztworu glikolu etylowego. Pułapki były eksponowane w terenie przez 5 miesięcy, od początku maja do końca września, a materiał wybierano z nich w odstępach miesięcznych. Zebrany materiał przesłano do Zakładu Zoologii Instytutu Biologii Uniwersytetu Humanistyczno-Przyrodniczego Jana Kochanowskiego w Kielcach, gdzie został poddany analizie faunistyczno-ekologicznej. Wyniki analiz przedstawiono w tabeli 61, a ich opracowanie stanowi odrębną część Raportu, wykonaną przez Uniwersytet Humanistyczno-Przyrodniczy Jana Kochanowskiego w Kielcach.
Tabela 61. Charakterystyka gatunków biegaczowatych odłowionych w 2012 roku
Tabela 62. Charakterystyka zgrupowania biegaczowatych odłowionych w 2012 roku
W borze bagiennym odłowiono 218 osobników należących do 10 gatunków (Tab. 61, 62). Wskaźnik łowności miał niską wartość wnoszącą 0,096 ale był ponad czterokrotnie wyższy niż w roku poprzednim. W strukturze troficznej przeważały zdecydowanie zoofagi duże, które stanowiły ponad 95% osobników zgrupowania, pozostałą część zgrupowania stanowiły osobniki gatunków należących do zoofagów małych. Nie stwierdzono hemizoofagów. Różnorodność była niska i wynosiła nieco ponad 38% wartości maksymalnej, co wynikało z dużej dominacji Pterostichus niger w badanym środowisku, który stanowił ponad 69% osobników zgrupowania. Łowność, chociaż wyższa niż w poprzednim roku, miała niską wartość. Ale w przypadku tego środowiska łowność Carabidae jest zawsze niska. Zdecydowana przewaga zoofagów dużych może wskazywać na dobry stan badanego środowiska. W lesie mieszanym odłowiono 1213 osobników, które należały do 10 gatunków (Tab. 61, 62). Łowność wyniosła 0,54 i była ponad trzykrotnie wyższa niż w roku 2011. W strukturze troficznej zdecydowanie dominowały osobniki zoofagów dużych (ponad 97%), pozostałą część zgrupowania stanowiły zoofagi małe, nie stwierdzono natomiast hemizoofagów. Wskaźnik różnorodności nie był wysoki – osiągnął ponad 38% wartości maksymalnej. Wskaźnik równomierności również kształtował się na niskim poziomie. Obie wartości wynikają ze zdominowania zgrupowania przez osobniki Carabus hortensis oraz Pterostichus niger, które stanowiły ponad 90% osobników zgrupowania.
5. Programy specjalistyczne
Stacja Bazowa WIGRY, oprócz podstawowych programów pomiarowych ZMŚP, realizuje również inne programy specjalistyczne. Programy te prowadzone są na obszarze zlewni badawczej ZMŚP lub jej otuliny - na terenie Wigierskiego Parku Narodowego.
5.1. Badania organizmów saproksylicznych w różnych ekosystemach leśnych
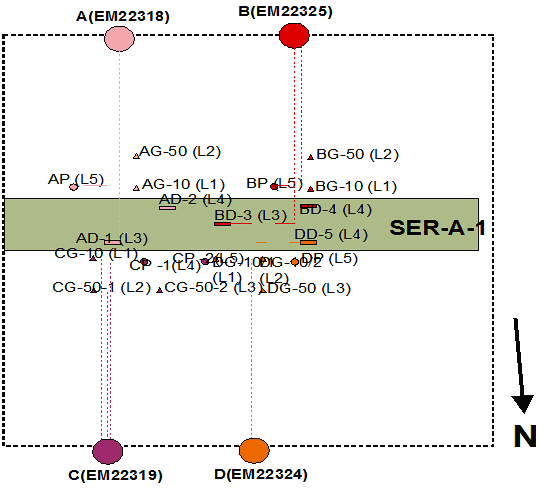
Badania polegają na określeniu składu gatunkowego wybranych grup organizmów saproksylicznych (wybranych gatunków owadów, śluzowców, mszaków oraz grzybów, w tym porostów), warunków ich występowania i preferencji siedliskowych oraz zagrożeń (Krzysztofiak red. 2012a). Organizmy saproksyliczne (zarówno grzyby, śluzowce, mszaki, jak i owady) odgrywają bardzo istotną rolę w prawidłowym funkcjonowaniu środowisk leśnych. Znajomość ich zasobów, rozprzestrzenienia oraz zagrożeń, jakim podlegają jest sprawą kluczową w ochronie tych organizmów oraz ochronie całych siedlisk przyrodniczych. Wiele gatunków tych organizmów objętych jest ochroną prawną oraz znajduje się na Czerwonych Listach. Wśród grzybów makroskopijnych 25% wszystkich gatunków stwierdzonych w Polsce umieszczonych zastało na Czerwonej Liście, w tym 71 gatunków ze statusem gatunku wymarłego i zaginionego, a 171 ze statusem gatunku wymierającego. Spośród śluzowców na 88 gatunków zamieszczonych na Czerwonej Liście aż 24 mają status gatunku zaginionego. Również wśród mszaków wiele jest gatunków zagrożonych wyginięciem - na Czerwonej Liście znalazło się około 20% wszystkich gatunków stwierdzonych w Polsce. Podobnie jest z owadami, do których zaliczamy również gatunki występujące w załącznikach Dyrektywy Siedliskowej. Głównym celem badań jest rozpoznanie różnorodności gatunkowej wybranych organizmów, zebranie danych dotyczących warunków ich występowania (preferencji siedliskowych) oraz określenie zagrożeń jakim podlegają. W przypadku grzybów szczegółowym przedmiotem badań są grzyby makroskopijne (macromycetes) oraz grzyby lichenizowane (porosty), związane z martwym drewnem. Do szczegółowych celów prowadzonych badań w tym zakresie należy: - zbadanie różnorodności grzybów, w tym porostów, związanych z martwym drewnem w wybranych zbiorowiskach leśnych; - oszacowanie spektrum bioekologicznych powiązań tych grzybów z zasiedlanym środowiskiem; - oszacowanie zagrożeń i możliwości przetrwania badanej grupy w warunkach lokalnych; - wskazanie kategorii, typów mikro- i makrosiedlisk oraz składników dendroflory ważkich z punktu widzenia utrzymania i/lub zwiększenia różnorodności biologicznej analizowanej grupy grzybów; - charakterystyka wybranych aspektów zmian czasowych mikobioty; - analiza powiązań procesu owocnikowania grzybów z wybranymi elementami meteorologicznymi. Szczegółowym celem badań śluzowców jest prześledzenie zmian ilościowych i jakościowych, jakim w ciągu możliwie długiego przedziału czasowego podlega ta grupa organizmów. Dzięki temu możliwe będzie poznanie fenologii śluzowców, określenie ich preferencji odnośnie podłoży oraz oszacowanie trwałości „owocowań”. Badania pozwolą prześledzić fenologię śluzowców w powiązaniu z zachodzącymi zmianami środowiskowymi. Obserwacje na stałych powierzchniach, na których czynniki środowiskowe są stale monitorowane, umożliwią znaczne poszerzenie wiedzy o ekologii śluzowców. W przypadku owadów szczegółowym przedmiotem opracowania są chrząszcze związane z martwym drewnem. Do szczegółowych celów prowadzonych badań w tym zakresie należy poznanie składu gatunkowego i struktury zgrupowań tych owadów w różnych ekosystemach leśnych. Spośród mszaków badaniami objęto przedstawicieli gromad Marchantiophyta i Bryophyta, występujących na rozkładającym się drewnie. Głównym celem badań jest rozpoznanie różnorodności gatunkowej mchów i wątrobowców porastających różne formy rozkładającego się drewna oraz określenie preferencji w stosunku do typu podłoża i zbiorowiska leśnego. Do określenia warunków meteorologicznych w badanych typach lasu (w grądzie, świerczynie bagiennej, sosnowo-brzozowym lesie bagiennym) służą trzy kompaktowe stacje meteorologiczne. Każda ze stacji dokonuje ciągłego pomiaru następujących parametrów: prędkość wiatru, kierunek wiatru, wielkość opadu atmosferycznego, wilgotność względna powietrza, temperatura powietrza, promieniowanie słoneczne całkowite. Na jednej powierzchni w grądzie wybrano do badań szczegółowych kłodę próchniejącego drewna, na której zainstalowano zestaw czujników mierzących temperaturę (powietrza - tuż przy kłodzie drewna, w kłodzie drewna oraz w glebie tuż przy kłodzie), wilgotność gleby, drewna i powietrza oraz przewodność elektrolityczną gleby i drewna. Łącznie zainstalowano na powierzchni 20 czujników: 5 w drewnie - w powierzchniowej warstwie, 5 w glebie - w odległości 10 i 50 cm od kłody, na głębokości 5 cm i 10 nad powierzchnią gleby - 10 cm od kłody i 10 cm nad powierzchnią gruntu (Rys. 101).
Rys. 101. Rozmieszczenie czujników i rejestratorów na powierzchni w grądzie. SER-A-1 - kłoda drewna; A(EM22318) ...
5.1.1. Powierzchnie badawcze
Badania prowadzone są na terenie Wigierskiego Parku Narodowego, na trzech stanowiskach, w trzech typach lasów: w grądzie, sosnowo-brzozowym lesie bagiennym oraz w świerczynie bagiennej. Na każdym stanowisku wyznaczonych zostało po 12 powierzchni badawczych o rozmiarach 5 x 5 m. Stanowisko w grądzie Tilio-Carpinetum zostało zlokalizowane w południowo-wschodniej części Parku, w oddziale 361. Na tym stanowisku zainstalowana została stacja meteorologiczna oraz zestaw czujników mierzących wybrane parametru kłody drewna i w jego najbliższego otoczenia. Stanowisko w sosnowo-brzozowym lesie bagiennym Thelypteridi-Betuletum pubescentis zlokalizowane zostało w północnej części Parku, w pobliżu doliny Wiatrołuży, w oddziale 20. Na tym stanowisku zainstalowana została stacja meteorologiczna, wokół której rozmieszczone zostały powierzchnie badawcze. Stanowisko w świerczynie bagiennej Sphagno girgensohnii-Piceetum zlokalizowane zostało w środkowej części Parku, w pobliżu doliny Czarnej Hańczy, w oddziale 121. Podobnie jak na poprzednim stanowisku zainstalowano tu stację meteorologiczną, a wokół niej wyznaczono 12 powierzchni badawczych.
5.1.2. Wyniki badań
5.1.2.1. Inwentaryzacja organizmów saproksylicznych
Śluzowce W badaniach prowadzonych w 2012 roku na trzech stanowiskach badawczych - w grądzie (Sernetki - SER), w sosnowo-brzozowym lesie bagiennym (Wiatrołuża - WIAT) i w świerczynie bagiennej (Dolina Czarnej Hańczy - HAN) stwierdzono występowanie 25 gatunków śluzowców. Są to: 1. Arcyria cinerea – strzępek wyprostowany – HAN (Fot. 1). 2. Arcyria denudata – strzępek błyszczący – SER. 3. Arcyria ferruginea – strzępek ciemny – HAN (Fot. 2). 4. Ceratiomyxa fruticulosa – śluzek krzaczkowaty – WIA, HAN, SER (Fot. 3). 5. Cribraria argillacea – przetaczek gromadny – HAN. 6. Cribraria vulgaris – przetaczek zmienny – SER). 7. Diderma ochraceum – szaroń brunatnawy – HAN. 8. Didymium iridis – makulec pokrewny – WIA (Fot. 4). 9. Fuligo septica – wykwit zmienny – HAN, SER. 10. Hemitrichia clavata – zapletka maczugowata – WIA, SER. 11. Hemitrichia serpula – zapletka czołgaczek – WIA, SER). 12. Lamproderma arcyrionema – błyszczek siatkowaty – WIA. 13. Lepidoderma tigrinum – łuskowiec tygrysowaty – SER, HAN. 14. Lycogala conicum – rulik stożkowaty – SER (Fot. 5). 15. Lycogala epidendrum – rulik groniasty – WIA, HAN. 16. Physarum album – maworek zwisły – WIA, HAN. 17. Reticularia lycoperdon – samotek zmienny – SER. 18. Stemonitis axifera – paździorek rdzawy – WIA, SER (Fot. 6). 19. Stemonitis fusca – paździorek ciemny – WIA, HAN, SER (Fot. 7). 20. Stemonitopsis typhina – paździoreczek gęsty – SER. 21. Trichia decipiens – kędziorek mylny – WIA, HAN, SER. 22. Trichia favoginea – kędziorek złocik – SER (Fot. 8). 23. Trichia scabra – kędziorek szorstki – WIA (Fot. 9). 24. Trichia varia – kędziorek różnokształtny – SER (Fot. 10). 25. Tubulifera arachnoidea – zlepniczek walcowaty – HAN (Fot. 11).
Fot. 1. Arcyria cinerea - strzępek wyprostowany
Najczęściej spotykanym gatunkiem był kędziorek mylny (Trichia decipiens), notowany 11 razy, na różnych podłożach. Sześciokrotnie znaleziono Physarum album i Stemonitis fusca, czterokrotnie Lycogala epidendrum. Są to wszystko gatunki pospolite. Znaleziono też jeden gatunek nowy dla Wigierskiego Parku Narodowego – Diderma ocheaceum.
Owady W trakcie badań odłowiono łącznie 1156 osobników chrząszczy należących do 14 rodzin i 34 gatunków. Najliczniej odławianymi gatunkami były:Cis glabratus (61,16% osobników), Cis jacquemartii (15,14%), Bolitophagus reticulatus (4,5%), Cis quadridens (2,94%), Ennearthron cornutum (2,94%), Cis boleti (2,25%), Cercyon convexiusculus (1,38%), Cyllodes ater (1,12%) i Cis micans (1,12%). Pozostałe gatunki osiągnęły udział procentowy poniżej 1%. Do klasy wierności grupującej owady obligatoryjnie związane z silnie rozłożonym drewnem i próchnowiskami (F3) należało 5,88% gatunków i 1,3% osobników. Owady związane ze środowiskiem podkorowym, owocnikami grzybów nadrzewnych i dziuplami (F2) stanowiły 79,41% gatunków i 96,63% odłowionych osobników. Do chrząszczy fakultatywnie związanych z zamierającymi drzewami lub rozkładającym się drewnem (F1) należało 2,94% gatunków i 0,09% osobników. Klasa wierności tworzona przez owady nie związane z rozkładającym się drewnem (F0) obejmowała 11,76% gatunków i 1,99% osobników. Najliczniejszą grupą troficzną były chrząszcze odżywiające się grzybami, stanowiły one 95,59% odłowionych osobników i 61,76% gatunków. Następne pod względem liczby odłowionych osobników były myksomykofagi, których udział wyniósł 1,73% osobników i 11,76% gatunków. Najmniej liczne okazały się chrząszcze drapieżne osiągając udział 0,95% osobników i 20,59% gatunków. Pozostałe 1,73% osobników i 5,88% gatunków stanowią chrząszcze o niedostatecznie poznanych preferencjach pokarmowych. Kompletna lista odłowionych gatunków wraz z klasami wierności i rzadkości faunistycznej oraz grupami troficznymi przedstawia tabela 63.
Tabela 63. Chrząszcze (Coleoptera) odłowione na badanych powierzchniach w 2012 r.
F – klasa wierności w stosunku do silnie rozłożonego drewna i próchnowisk: 3 – gatunki obligatoryjnie związane z silnie rozłożonym drewnem i próchnowiskami; 2 – gatunki mniej związane z samym stanem rozkładu drewna, lecz preferujące owocniki grzybów nadrzewnych, środowisko podkorowe, dziuple itp.; 1 – gatunki fakultatywnie związane z zamierającymi drzewami lub rozkładającym się drewnem, często zasiedlające leżaninę i drzewa w różnym stopniu osłabienia; 0 – gatunki nie związane z rozkładającym się drewnem; T – grupa troficzna: M – mykofagi, MX – Myksomykofagi, Z – zoofagi, ? – gatunki o niedostatecznie poznanych preferencjach pokarmowych; R – gatunki należące do rzadkości faunistycznych lub/i reliktów lasów pierwotnych; N – gatunki nowe dla Pojezierza Mazurskiego
Grzyby W trakcie przeprowadzonych badań stwierdzono łącznie występowanie 85 taksonów grzybów makroskopijnych związanych z martwym drewnem, z czego 76 taksonów stanowią podstawczaki, a 9 - przedstawiciele workowców.
Na badanych powierzchniach na murszejącym drewnie znaleziono 40 taksonów porostów (Tab. 64). Spektrum morfologiczne znalezionych porostów przedstawia się następująco: skorupiaste – 20 taksonów; łuseczkowate – 1; listkowate – 9; krzaczkowate – 3; wielopostaciowe – 7.
Tabela 64. Częstość występowania taksonów porostów w badanych środowiskach
Spektrum biologiczne jest typowe dla porostów analizowanego podłoża i badanych siedlisk leśnych. Pod względem liczby gatunków oraz częstości i obfitości występowania przeważają porosty humusolubne i cienioznośne (rodzaje Cladonia, Placynthiella, Trapeliopsis, Dimerella, Micarea, Lepraria – razem 16 taksonów). Większość stanowią porosty leśne, a gatunków typowo światłolubnych i charakterystycznych dla środowisk nieleśnych prawie nie ma. Wyjątek stanowi Ramalina farinacea, która jest porostem z równą częstotliwością rosnącym w lasach i poza nimi, ale jednocześnie jest gatunkiem światłolubnym, w lasach rosnącym głównie wysoko na pniach drzew lub w ich koronach. Zdecydowanie największe zróżnicowanie epiksylicznych porostów stwierdzono w grądzie, gdzie znaleziono ponad dwa razy więcej taksonów niż w obu pozostałych typach lasów. Znacznie mniejsza niż w grądzie liczba taksonów epiksyli (po 16) w borealnej świerczynie bagiennej i w brzozowo-sosnowym lesie bagiennym, wynika z ubóstwa bioty porostów epifitycznych w tych zbiorowiskach. Kora sosen, świerków i brzóz, które tworzą drzewostany w tych zbiorowiskach, stwarza porostom bardzo niekorzystne warunki do rozwoju. Poza kilkoma gatunkami sporadycznymi nie stwierdzono taksonów wyróżniających oba te zbiorowiska.
Mszaki W obrębie analizowanych 36 powierzchni, na siedliskach murszejącego drewna, stwierdzono występowanie 76 taksonów mszaków, tj. 51 gatunków i jednej odmiany mchów oraz 24 gatunków wątrobowców. Oznacza to, że na badanych podłożach występuje 34% mszaków znanych z całego Wigierskiego Parku Narodowego. W przypadku wątrobowców, liczba ta jest jeszcze wyższa i wynosi ona aż 55% wszystkich przedstawicieli hepatikoflory Parku. Liczebnie dominują taksony występujące rzadko – 54%, częstych jest 28%, a pospolitych – 18%. Do najczęściej notowanych gatunków należą: mchy – Brachythecium salebrosum, Dicranum scoparium (Fot. 26), Eurhynchium angustirete (Fot. 27), Herzogiella seligeri (Fot. 28), Hylocomium splendens (Fot. 29), Hypnum cupressiforme (Fot. 30), Hypnum pallescens (Fot. 31), Orthodicranum montanum, Plagiothecium laetum, Pleurozium schreberi, Sanionia uncinata i Tetraphis pellucida oraz dwa wątrobowce – Lepidozia reptans, Lophocolea heterophylla (Fot. 32).
Stwierdzone w trakcie badań taksony należą do 47 rodzajów i 28 rodzin. Najliczniejszymi rodzinami są: wśród mchów – Hypnaceae (8 gat.), Brachytheciaceae (7), Dicranaceae (5), Sphagnaceae (8), a w przypadku wątrobowców – Calypogeiaceae (4), Cephaloziaceae (4), Geocalycaceae. Gatunki odnotowane na próchniejących obiektach mają bardzo różny charakter ekologiczny. W pierwszych stopniach rozkładu na korze wciąż utrzymują się epifity takie jak np. Radula complanata czy Metzgeria furcata. Wraz z postępującym stopniem rozkładu pojawiają się tzw. właściwe epiksyle (gatunki stricte związane z rozkładającym się drewnem). Do takich zaliczane są głównie wątrobowce: Blepharostoma trichophyllum,Riccardia latifrons, Nowellia curvifolia i Geocalyx graveolens, ale także i niektóre mchy np. Aulacomniu androgynum, Callicladium haldanianum, Herzogiella seligeri czy Tetraphis pellucida. W ostatnich fazach dekompozycji na drewnie pojawiają się gatunki przechodzące z dna lasu. Są to m.in. Calliergonella cuspidata, Climacium dendroides, Dicranum polystetum, Pleurozium schreberi, Rhytidiadelphus triquetrus oraz wszystkie gatunki z rodzaju Sphagnum. Na podłożach rozkładającego się drewna w badanych fitocenozach leśnych stwierdzono występowanie 25 gatunków mszaków objętych ochroną (8 ochroną ścisłą, 17 ochroną częściową), które stanowią łącznie ponad 30% analizowanej musko- i hepatikoflory. Na badanych typach siedlisk zanotowano także obecność trzech zagrożonych gatunków wątrobowców – Nowellia curvifolia (Fot. 33), Calypogeia suecica i Geocalyx graveolens (Fot. 34).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||