|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
4.10. Fauna epigeiczna
Badania dotyczące fauny epigeicznej dotyczyły tylko chrząszczy z rodziny biegaczowatych Carabidae i prowadzone były w dwóch typach siedliskowych lasu - w borze bagiennym i lesie mieszanym. Na każdym siedlisku wytypowano po trzy stanowiska oddalone od siebie o około 250 m, na których zainstalowano po 5 pułapek rozmieszczonych liniowo co trzy metry. Pułapki (słoiki o pojemności 0,33 l i średnicy otworu 56 mm) zostały wkopane równo z powierzchnią gruntu i osłonięte daszkami. Jako płynu konserwującego użyto roztworu glikolu etylowego. Pułapki były eksponowane w terenie przez 5 miesięcy, od początku maja do końca września, a materiał wybierano z nich w odstępach miesięcznych. Analizę zebranych biegaczowatych przeprowadzono w Zakładzie Zoologii Instytutu Biologii Uniwersytetu Humanistyczno-Przyrodniczego Jana Kochanowskiego w Kielcach. Do oceny wpływu czynników środowiskowych na faunę biegaczowatych posłużono się odpowiednimi wskaźnikami ekologicznymi:
Struktura dominacji zgrupowania. Ustalamy ją, określając dominację indywidualną każdego gatunku za pomocą wzoru
gdzie: n – liczba osobników danego gatunku, N – liczba wszystkich osobników badanej grupy taksonomicznej. W zależności od procentowego udziału danego gatunku w zgrupowaniu, zaliczamy go do określonej klasy dominacji. Przyjęto podział wskaźnika dominacji na 5 klas (Górny, Grüm 1981): D1 – subrecedenty: gatunki, których osobniki stanowią ≤ 1,0% osobników zgrupowania; D2 – recedenty: gatunki, których osobniki stanowią 1,1-2,0% osobników zgrupowania; D3 – subdominanty: gatunki, których osobniki stanowią 2,1-5,0% osobników zgrupowania; D4 – dominanty: gatunki, których osobniki stanowią 5,1-10,0% osobników zgrupowania; D5 – eudominanty: gatunki, których osobniki stanowią > 10,0% osobników zgrupowania.
Różnorodność zgrupowań. Różnorodność zgrupowań oszacowana została przy pomocy wskaźnika różnorodności Shanonna i Weavera (H’)
gdzie
pi oznacza frakcję
Równomierność podziału osobników między gatunki. Oceniona została za pomocą wskaźnika równomierności Shannona-Weavera (J’)
gdzie H’ – wartość wskaźnika różnorodności Shanonna i Weavera, S – liczba gatunków Analizę zoogeograficzną zebranego materiału dokonano w oparciu o klasyfikację Leśniaka (1987).
4.10.1. Bór bagienny
Na powierzchni zlokalizowanej w borze bagiennym odłowiono 53 chrząszcze z rodziny biegaczowatych Carabidae, które należały do 5 gatunków (Tab. 58). W odłowionym materiale zabrakło chrząszczy z gatunku Pterostichus nigrita, które w poprzednim roku był odławiane na tej powierzchni. Średnia łowność biegaczowatych w pułapki Barbera wynosiła 0,02 osobnika na pułapkę w ciągu doby. W odłowionym materiale jako eudominaty występowały dwa gatunki - szykoń czarny Pterostichus niger i biegacz fioletowy Carabus violaceus. Szykoń czarny jest leśnym gatunkiem „wszędobylskim” – występuje zarówno w drzewostanach liściastych jak i iglastych. Jego najwyższa aktywność przypadała na lipiec. Biegacz fioletowy, tak jak i pozostałe gatunki z tego rodzaju, objęty jest ochroną ścisłą (Rozporządzenie Ministra Środowiska, 2004). Jest to szeroko rozprzestrzeniony gatunek europejski, najczęściej spotykany w lasach liściastych i mieszanych.
Tabela 58. Charakterystyka gatunków biegaczowatych odłowionych w borze bagiennym w 2010 roku
Struktura dominacji w zgrupowaniu biegaczowatych przedstawia się następująco: - eudominanty (D5) stanowiły 83,02%; - dominanty (D4) stanowiły 15,09% - recedenty (D2) stanowiły 1,89%. W strukturze troficznej zdecydowanie dominowały duże zoofagi (Tab. 59), osobniki o ciężarze powyżej 100 mg, odżywiające się głównie pokarmem zwierzęcym. Stanowiły one 91,8% wszystkich odłowionych na powierzchni biegaczowatych. Ponadto występowały małe zoofagi, osobniki odżywiające się również pokarmem zwierzęcym ale o wadze poniżej 100 mg. Stanowiły one 8,2% odłowionych biegaczowatych.
Tabela 59. Struktura troficzna zgrupowania biegaczowatych w borze bagiennym dla dostępnych lat obserwacji
Tabela 60. Charakterystyka zgrupowania biegaczowatych boru bagiennego dla dostępnych lat obserwacji
Zgrupowanie biegaczowatych występujące w borze bagiennym jest dość ubogie – stwierdzono zaledwie 5 gatunków. Wskaźnik łowności kształtował się na bardzo niskim poziomie i wynosił 0,02 osobnika na pułapkę w ciągu doby (Tab. 60). Natomiast wysokie wartości osiągnęły wskaźniki struktury troficznej, wskaźnik różnorodności oraz równomierności. Struktura troficzna była zdominowana przez zoofagi duże (91,8%), przy jednoczesnym braku w zgrupowaniu Carabidae hemizoofagów. Wskaźnik różnorodności osiągnął ponad 73% wartości maksymalnej, a wskaźnik równomierności prawie 66% wartości maksymalnej. Wartości wszystkich wskaźników wskazują na dobry stan badanego środowiska, a mała łowność chrząszczy jest charakterystyczna dla tego typu środowiska.
4.10.2. Las mieszany
W lesie mieszanym odłowiono łącznie 354 chrząszcze z rodziny Carabidae, należące do 9 gatunków (Tab. 61). W porównaniu do poprzedniego roku w odłowionym materiale nie stwierdzono tylko jednego gatunku - Carabus granulatus. Pojawiły się natomiast dwa inne: Carabus auronitens i Nebria brevicollis. Średnia łowność biegaczowatych w pułapki Barbera wynosiła 0,16 osobnika na pułapkę w ciągu doby i była ośmiokrotnie wyższa, niż w borze bagiennym. Rolę eudominanta w odłowionym materiale odgrywały trzy gatunki - szykoń czarny Pterostichus niger, tak, jak w poprzednim roku, biegacz ogrodowy Carabus hortensis oraz Pterostichus melanarius. Szykoń czarny stanowił 58,5% wszystkich odłowionych biegaczowatych. Biegacz ogrodowy odławiany był znacznie rzadziej i stanowił ponad 16% wszystkich odłowionych biegaczowatych, podobnie jak Pterostichus melanarius, który stanowił ponad 12% całego materiału. Struktura dominacji w zgrupowaniu przedstawia się następująco: - eudominanty (D5) stanowiły 87,0%; - dominanty (D4) stanowiły 6,5% - subdominant (D3) stanowiły 3,4%, - recendenty (D2) stanowiły 1,4%, - subrecedenty (D1) stanowiły 1,7%.
Tabela 61. Charakterystyka gatunków biegaczowatych odłowionych w lesie mieszanym w 2010 roku
W strukturze troficznej dominowały, tak, jak i w borze bagiennym, duże zoofagi (Tab. 61, 62). Stanowiły one 92,6% wszystkich odłowionych na powierzchni biegaczowatych. Ponadto występowały małe zoofagi, które stanowiły 7,4% odłowionych biegaczowatych. W lesie mieszanym stwierdzono 8 gatunków biegaczowatych. Wielkość wskaźnika różnorodności Shannona-Weavera wynosi H’=2,079 a równomierności J’=0,313 (Tab. 63).
Tabela 62. Struktura troficzna zgrupowania biegaczowatych w lesie mieszanym dla dostępnych lat obserwacji
Tabela 63. Charakterystyka zgrupowania biegaczowatych lasu mieszanego dla dostępnych lat obserwacji
Zgrupowanie biegaczowatych występujące w lesie mieszanym jest stosunkowo ubogie - odłowiono jedynie 354 osobniki z 9 gatunków. Wielkość łowności uznać można w tym środowisku za zbyt niską, a tym samym wskazującą na istnienie zakłóceń w środowisku. Jednak nie potwierdzają tego wartości innych wskaźników. W strukturze troficznej zdecydowanie dominowały zoofagi duże (ponad 92%), przy jednoczesnym braku hemizoofagów. Wskaźniki różnorodności i równomierności osiągnęły ponad 59% wartości maksymalnej.
Literatura Górny M., Grüm L. 1981. Metody stosowane w zoologii gleby. PWN, Warszawa, s. 483. Leśniak A. 1987. Zoogeographical analysis of the Carabidae (Coleoptera) of Poland. Fragm. Faun., Warszawa, 30, s. 297-312.
5. Ocena stanu środowiska i zagrożenia
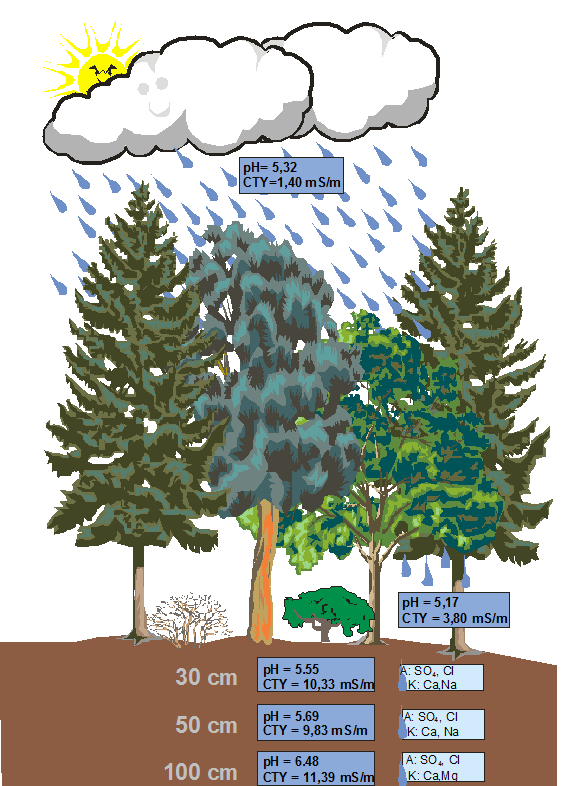
Rok 2010 pod wieloma względami różnił się od poprzednich lat badań, a zwłaszcza ostatniego roku. Nie zarejestrowano w tym okresie, podobnie jak w poprzednio, zjawisk drastycznie pogarszających stan środowiska zlewni badawczej WIGRY. Zdecydowana większość badanych parametrów fizyczno-chemicznych środowiska była na podobnym poziomie, co w latach poprzednich. Obserwacje przyrodnicze przeprowadzone w 2010 roku ponownie potwierdziły, że stan przyrody zlewni i jej otoczenia w bardzo dużym stopniu zależy od warunków termiczno-opadowych. To temperatura powietrza, gleby i wody decyduje o przeżywalności i aktywności wielu gatunków roślin i zwierząt, a dostępność wody dla tych organizmów, zwłaszcza na początku sezonu wegetacyjnego, decyduje o ich liczebności, sukcesie rozrodczym i kondycji. Miniony rok pod względem termiczno-opadowym został oceniony jako normalny-wilgotny i znacznie odbiegał od poprzednich lat badań (Rys. 3). Maksymalne temperatury powietrza przekraczały 30°C i rejestrowane były w czerwcu, lipcu i sierpniu (Tab. 4). Najcieplejszym miesiącem był lipiec, ze średnią temperaturą 21,21°C, a najchłodniejszym styczeń, ze średnią temperaturą -10,59°C. Najniższa temperatura powietrza, jaką zarejestrowano w ciągu roku wynosiła -29,10°C (styczeń). Rok kalendarzowy 2010 należał do lat najchłodniejszych w ciągu całego okresu badań, a średnia roczna temperatura wynosiła 6,34°C (chłodniejszy był tylko rok poprzedni). Była to wartość niemal identyczna z temperaturą średnią z wielolecia 1971-2001. W ciągu roku wystąpiło 87 dni z temperaturą maksymalną poniżej zera, a 15 dni z temperaturą maksymalną powyżej 30°C. W kwietniu wystąpił okres przymrozkowy trwający przez 10 dni, ze średnią dobową temperaturą -4,2°C (minimalne temperatury dochodziły do ponad -7°C). Pomimo tego średnia miesięczna temperatura kwietnia wyniosła 7,33°C. Różnica między najcieplejszym i najchłodniejszym dniem wynosiła ponad 28°C. W ciągu roku wystąpiły trzykrotnie gwałtowne skoki średniej dobowej temperatury ponad 10°C - w styczniu (-11,28°C ), w maju (-10,19°C ) i we wrześniu (12,38°C ). W minionym roku wystąpiła też jedna noc tropikalna (24 sierpnia), z temperaturą minimalną 20,2°C (maksymalna temperatura dochodziła prawie do 25°C, a średnia temperatura wyniosła 21,69°C). Biorąc pod uwagę pomiary temperatury powietrza z całego okresu badań (2002-2010) wyznaczono termiczne pory roku. Najdłuższą porą jest zima, która trwa tu średnio 102 dni. Po niej następuje krótkie przedwiośnie, trwające 29 dni, które przechodzi w wiosnę (55 dni). Jesień trwa średnio 60 dni i z reguły łagodnie przechodzi w przedzimie, które zwykle trwa 31 dni. Suma roczna opadów atmosferycznych w minionym roku wyniosła 715,2 mm i należała do najwyższych od czasu prowadzenia obserwacji, czyli od 2002 roku. Była ona wyższa o 73,8 mm od sumy opadów z poprzedniego roku i aż o 124,0 mm od średniej z wielolecia. Najbardziej odbiegały od średniej z wielolecia wielkości opadów z maja - były prawie o 90% wyższe (Rys. 10). Najwięcej opadów wystąpiło w miesiącu maju - 138,8 mm, co stanowiło ponad 19% rocznych opadów oraz w sierpniu - 109,0 mm (ponad 15%). Opady w maju występowały przez 20 dni, a maksymalny opad dobowy wynosił 23,2 mm, natomiast opady w sierpniu trwały 12 dni, z tym, że maksymalny opad dobowy dochodził nawet do 43,2 mm. Najmniej opadów zanotowano w styczniu - w ciągu 10 dni spadło 19,8 mm opadów. W tym miesiącu zanotowano również najdłuższy okres bezopadowy - trwał on 15 dni, od 13 do 27 stycznia. Ogólnie można stwierdzić, że warunki termiczno-opadowe w minionym roku były sprzyjające dla przyrody. Zwłaszcza dość wysokie opady atmosferyczne znacząco i przez dłuższy okres zasilały liczne bezodpływowe zagłębienia terenu, w których bujnie rozwijały się specyficzne zespoły roślin i zwierząt. Okres wegetacyjny rozpoczął się już 19 marca i trwał do 14 października, czyli 210 dni. Na stan przyrody duże znaczenie ma nie tylko ilość opadów atmosferycznych, ale również jakość tych opadów, będąca pośrednim wskaźnikiem stopnia zanieczyszczenia powietrza atmosferycznego. W 2010 roku jakość opadów nie odbiegała zasadniczo od poziomu stwierdzonego w poprzednim roku. Na jeden km 2 niezalesionej powierzchni zlewni badawczej spadło z opadami atmosferycznymi 2368,55 kg głównych jonów, czyli o ponad 42% mniej, niż w poprzednim roku. Przeważały jony potasowe i amonowe. Stwierdzone stężenia poszczególnych jonów w opadach atmosferycznych wskazują, że ogólnie można je sklasyfikować jako normalne pod względem odczynu i o nieznacznej przewodności elektrolitycznej. Można zatem pośrednio wnioskować, że powietrze w rejonie zlewni badawczej nie jest zanieczyszczone. Na obszarach zalesionych zlewni badawczej (z drzewostanem sosnowo-świerkowym) opad atmosferyczny przechodząc przez warstwę drzew ulegał znacznym modyfikacjom i to zarówno pod względem ilościowym, jak i jakościowym. W minionym roku hydrologicznym drzewostan zatrzymał średnio 56,35% opadów atmosferycznych. W poszczególnych miesiącach intercepcja podokapowa wahała się od 11,60% (luty) do 90,60% (wrzesień) wód opadowych. Wysoka intercepcja opadów w niektórych miesiącach powodowała, że nawet przy przeciętnych opadach gleba leśna była często przesuszona, co oczywiście wpływało ujemnie na rozwój roślin i niektórych zwierząt bezkręgowych. Pod względem jakościowym najistotniejsze zmiany dotyczą odczynu wody opadowej. Drzewostan sosnowo-świerkowy, a zwłaszcza świerki, powodują zakwaszenie wód. W minionym roku hydrologicznym średnia wartość pH opadów pod okapem drzew wynosiła 5,17 i była niemal identyczna, jak w roku ubiegłym. Najniższe wartości pH odnotowano w lutym (4,41), najwyższe zaś w wodach opadowych ze stycznia (7,34). Na obszarach zalesionych depozycja głównych jonów do podłoża była prawie o 500 kg/km2 większa, niż na terenach otwartych. Na 1 km 2 zalesionej powierzchni spadło 2868,2 kg głównych jonów, czyli o 1198,1 kg mniej, niż w poprzednim roku. Przy tak niewielkich stężeniach jonów trudno przypuszczać, aby ich dopływ do gleby z opadem atmosferycznym stanowił istotne zagrożenie dla prawidłowego funkcjonowania środowiska przyrodniczego zlewni. Wyniki badań wód opadowych, opadu podkoronowego i roztworów glebowych ukazują pewne procesy zachodzące w środowisku. Wody opadowe docierające do koron drzew charakteryzowały się normalnym odczynem - pH 5,32 i nieznacznej przewodności elektrolitycznej - 1,40 mSm (Rys. 81). Po przejściu przez warstwę drzew odczyn opadu był nadal normalny - średnia wartość pH tylko nieznacznie obniżyła się (do 5,17), a przewodność była znacznie podwyższona - wzrosła o 63% (do 3,80 mSm). Wody przechodzące przez korony drzew spłukują pył zgromadzony na powierzchni liści, zawierający liczne pierwiastki oraz wymywają niektóre jony z liści. Pewne pierwiastki są absorbowane z opadów przez korony drzew. Zjawisko to jest bardzo złożone i zależy od wielu czynników, m.in. od gatunku drzewa, powierzchni aparatu asymilacyjnego, trofii siedliska, reakcji jonowymiennych czy wielkości opadów atmosferycznych i ich zanieczyszczenia. W skali całego minionego roku tylko jony amonowe były zatrzymywane w koronach drzew, a pozostałe jony (siarczanowe, chlorkowe, azotanowe, magnezowe, sodowe, potasowe, chlorkowe i wapniowe) były wydalane. Absorpcja lub wydalanie jonów przez roślinność może być wynikiem reakcji wymiennych, np. zaobserwowana absorpcja jonu amonowego może zmniejszyć realną efektywność absorbowania jonów sodowych i potasowych (Stachurski, Zimka 2004). Podobna zależność może zachodzić między jonami wapnia i potasu. Zdaniem Stachurskiego i Zimki (2004) reakcje jonowymienne mogą umożliwiać roślinności szybkie i bezpośrednie zdobywanie deficytowych biogenów oraz usuwanie z roślinności jonów najmniej pożądanych (np. Na i K). W świetle wyników innych badań tych autorów (1984) azot zawarty w opadach atmosferycznych jest silnie absorbowany w strefie koron drzew. W 2010 roku w przypadku azotu azotanowego nie stwierdzono takiej prawidłowości, natomiast w przypadku azotu amonowego średnio z opadem podkoronowym docierało do podłoża ponad dwukrotnie mniej amoniaku, niż docierało do koron drzew. Część pierwiastków biogennych jest wprowadzana do obiegu przez roślinność drzewiastą w postaci martwej materii organicznej (opad organiczny). W minionym roku na każdy metr kwadratowy lasu spadło 488,84 g s.m. opadu organicznego, głównie w postaci igieł (64,4%). W ten sposób każdy hektar lasu w ciągu roku wzbogacił się o 871 kg węgla organicznego, prawie 22 kg azotu ogólnego, ponad 17 kg wapnia, ponad 2 kg potasu, 2 kg magnezu i tyle samo siarki ogólnej oraz prawie 1,5 kg fosforu ogólnego. Wody przedostające się przez korony drzew docierają do dna lasu i przenikają w głąb profilu glebowego. W warstwie powierzchniowej (do 30 cm) nastąpił wzrost wartości pH roztworów o prawie 7% (do 5,55) oraz wzrost o ponad 60% ich mineralizacji (do 10,33 mS/m). Wielkość pH na głębokości 50 cm jeszcze nieznacznie wzrosła - do 5,69, a przewodność obniżyła się do 9,83 mS/m. Na głębokości 100 cm oba te parametry wzrosły - pH do 6,48 a przewodność do 11,39 mS/m. W roztworach dominowały kationy wapnia i sodu, a wśród anionów jony siarczanowe i chlorkowe (Rys. 81). Pomimo wysokich opadów atmosferycznych w minionym roku średni stan dobowy wody na Czarnej Hańczy wyniósł 228 cm (podobnie jak w roku poprzednim) i należał do najniższych w okresie prowadzenia pomiarów, tj. w latach 1997-2010. Nadal ponad połowa stanów (59%) mieściła się w najniższym przedziale, czyli 220-229 cm, a część stanów (4%) zawierała się w jeszcze niższym przedziale 210-219 cm.
Rys. 81. Modyfikacje wód opadowych na obszarach leśnych zlewni badawczej Stacji Bazowej WIGRY w 2010 roku
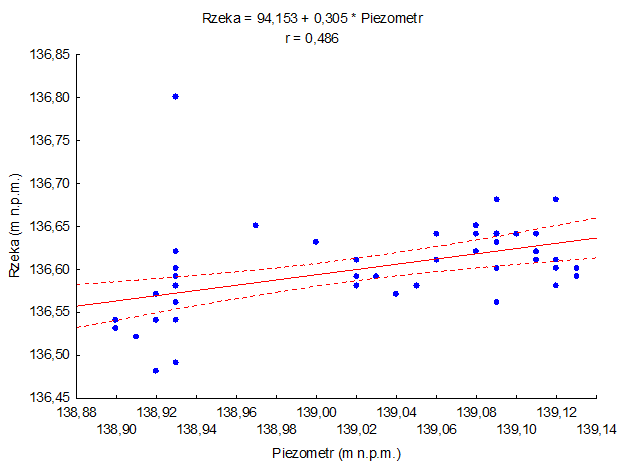
Najczęściej najniższe stany występowały w miesiącach zimowych, pod koniec lata i w okresie jesiennym (Rys. 57). Wysokie opady atmosferyczne w 2010 roku, wyższe prawie o 74 mm w porównaniu z poprzednim rokiem, spowodowały wzrost poziomu wód podziemnych. Średni miesięczny stan wód był wyższy o 5 cm od stanu z roku poprzedniego. Na koniec roku hydrologicznego poziom wód gruntowych był o 15 cm wyższy niż na początku roku, a maksymalna różnica między najniższym i najwyższym stanem dochodziła do 22 cm. Zagrożenie pojawienia się niżówki - suszy gruntowej istniało w całym roku hydrologicznym (Tab. 39). Badania właściwości fizyko-chemicznych wód podziemnych wskazują na ich wysoką jakość i świadczą o niewielkim oddziaływaniu lokalnych źródeł zanieczyszczenia. Wszystkie stężenia badanych jonów mieściły się w zakresie tła chemicznego, wyznaczonego dla obszaru zlewni na podstawie danych z wielolecia. Zgodnie z rozporządzaniem Ministra Środowiska z dnia 23 lipca 2008 r. w sprawie kryteriów i sposobu oceny stanu wód podziemnych określono stan chemiczny wody z piezometru w Sobolewie jako dobry. Średni roczny przepływ wód Czarnej Hańczy w Sobolewie był nieznacznie wyższy, niż w roku ubiegłym i wynosił 1,232 m3/s. Wyraźnie zaznaczyły się też różnice w przepływach miedzy półroczem zimowym i letnim - zimą SQ był niższy i wynosił 1,085 m3/s, a latem wzrósł do 1,379 m 3/s. W roku hydrologicznym 2010 przepłynęło w Sobolewie 38,9 mln m3 wody, czyli o 3,4 mln m3 więcej niż w roku poprzednim. Jakość tej wody (na podstawie rozporządzenie Ministra Środowiska z dnia 11 lutego 2004 roku w sprawie klasyfikacji dla prezentowania stanu wód powierzchniowych i podziemnych, sposobu prowadzenia monitoringu oraz sposobu interpretacji wyników i prezentacji stanu tych wód), podobnie jak i na Ujściu, odpowiadała III klasie czystości, czyli wodom o zadawalającej jakości, gdzie wartości biologicznych wskaźników wskazują umiarkowany wpływ oddziaływań antropogenicznych. Wody powierzchniowe są w określonym związku hydraulicznym z wodami podziemnymi. W odniesieniu do stanów wód Czarnej Hańczy położenie pierwszego zwierciadła wód podziemnych w piezometrze w Sobolewie kształtuje się powyżej stanów rzeki. W minionym roku, podobnie jak i w poprzednim, stwierdzono statystycznie istotną zależność pomiędzy stanami wód podziemnych, a stanami wód Czarnej Hańczy (Rys. 88).
Rys. 82. Zależność między poziomem wód w Czarnej Hańczy, a poziomem lustra wód podziemnych w Sobolewie (2010)
Na dobry stan środowiska zlewni badawczej składa się nie tylko wysoka jakość wód powierzchniowych i podziemnych, ale również niski poziom zanieczyszczeń w powietrzu atmosferycznym. Dla rok kalendarzowy zawartość dwutlenku siarki była niższa od progu detekcji. Zanieczyszczenie powietrza NO 2 było również niewielkie - 6,00 μg/m3. Wartość ta jest pięciokrotnie niższa od dopuszczalnego poziomu (traktowanego jako suma dwutlenku azotu i tlenku azotu w przeliczeniu na dwutlenek azotu), przyjętego dla ochrony zdrowia ludzi i roślin (rozporządzenie MŚ z dn. 3 marca 2008 r. w sprawie poziomów niektórych substancji w powietrzu. Dz. U. Nr 47, Poz. 281, z dnia 19 marca 2008 r.). Oba te związki są bardzo istotne z punktu widzenia ochrony środowiska, gdyż odpowiadają za zakwaszanie opadów atmosferycznych (Malzahn, 1999). Jednym z ważniejszych źródeł zanieczyszczenia powietrza są ciepłownie miejskie w Suwałkach, oddalone od granic zlewni o niecałe 10 km. Przy wiatrach wiejących z sektora północno-zachodniego wnoszą one nad teren zlewni pewne ilości dwutlenku azotu i dwutlenku siarki. Nie powoduje to jednak przekroczeń dopuszczalnych stężeń średniorocznych tych substancji w powietrzu, a tym samym nie zaburza funkcjonowania układów przyrodniczych na terenie zlewni, czy całego Wigierskiego Parku Narodowego. Potwierdzeniem tego są niskie stężenia i ładunki badanych jonów w wodzie opadowej, która wypłukuje zawarte w powietrzu zanieczyszczenia. Stan monitorowanych powierzchni porostów nadrzewnych był bardzo zbliżony do tego sprzed roku. Z 12 badanych powierzchni nadal na 5 nie stwierdzono monitorowanych gatunków porostów - ustąpiły one z tych powierzchni już w 2008 roku. Na pozostałych powierzchniach w 3 przypadkach zarejestrowano wyłącznie wzrost ogólnej powierzchni plech wszystkich monitorowanych gatunków, a w pozostałych przypadkach część gatunków zwiększyła powierzchnię plech, a część zmniejszyła. Gatunki porostów, które ustąpiły z monitorowanych powierzchni należą do porostów listkowatych (Hypogymnia physodes) oraz krzaczkowatych (Usnea hirta i Pseudevernia furfuracea). Badania zdrowotności drzew i drzewostanów wykazały, że w przypadku boru bagiennego drzewostan zaliczono do 2 klasy uszkodzeń (uszkodzenia lekkie), w przypadku świerczyny na torfie badany drzewostan zaliczono do 1 klasy uszkodzeń (stan ostrzegawczy), a w przypadku ubogiego grądu badany drzewostan zaliczono do 0 klasy uszkodzeń (brak uszkodzeń). W faunie biegaczowatych zaszły niewielkie zmiany w ilości gatunków występujących w badanych środowiskach. W borze bagiennym stwierdzono o jeden gatunek mniej niż w roku poprzednim, natomiast w lesie mieszanym zwiększyła się liczba gatunków biegaczowatych (o jeden gatunek). Na obu powierzchniach, podobnie jak w roku poprzednim, zdecydowanie dominował ten sam gatunek – Pterostichus niger, który stanowił około 50% wszystkich odłowionych biegaczowatych. Wartości wskaźników różnorodności i równomierności wskazują na dobry stan środowiska boru bagiennego, natomiast w lesie mieszanym mała łowność chrząszczy wskazuje na pewne zakłócenia w środowisku. Podsumowując uzyskane wyniki badań należy stwierdzić, że obszar zlewni badawczej Stacji Bazowej WIGRY charakteryzuje się dobrym stanem środowiska przyrodniczego. Zaobserwowane zmiany mają przeważnie charakter naturalny i na ogół nie wpływają negatywnie na stan środowiska. Rezultaty prowadzonego monitoringu mogą być bardzo przydatne dla oceny zachowania walorów przyrodniczych obszaru objętego najwyższą formą ochrony w Polsce - parku narodowego, Konwencją Ramsarską skupiającą najcenniejsze obszary wodno-błotne świata oraz Europejską Siecią Ekologiczną Natura 2000.
Literatura
Malzahn E., 1999: Ocena zagrożeń i zanieczyszczenia środowiska leśnego Puszczy Białowieskiej. Prace Inst. Bad. Leśn. Seria A, Warszawa. Stachurski A., Zimka J. R. 1984. The budżet of nitrogen dissolved in rainfall during its passing through the crown in forest ecosystems. Ekol. Pol. 32, s. 191-218. Stachurski A., Zimka J. R. 2004. Obieg pierwiastków w ekosystemach lądowych. Kosmos, 53, 1, s. 95-105. Stachurski A., Zimka J. R., Kwiecień M., 1994: Niektóre aspekty krążenia pierwiastków w ekosystemach leśnych na terenie Jakuszyc (II). (w:) Karkonoskie Badania Ekologiczne, II Konferencja, Dziekanów Leśny, 17-19.01.1994 (red. Fischer Z.), Oficyna Wydawnicza Instytutu Ekologii PAN, Dziekanów Leśny: 207-232.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||