|
4.10. Fauna epigeiczna
Badania dotyczące fauny epigeicznej dotyczyły tylko chrząszczy z rodziny biegaczowatych Carabidae i prowadzone były w dwóch typach siedliskowych lasu - w borze bagiennym i lesie mieszanym. Na każdym siedlisku wytypowano po trzy stanowiska oddalone od siebie o około 250 m, na których zainstalowano po 5 pułapek rozmieszczonych liniowo co trzy metry. Pułapki (słoiki o pojemności 0,33 l i średnicy otworu 56 mm) zostały wkopane równo z powierzchnią gruntu i osłonięte daszkami. Jako płynu konserwującego użyto roztworu glikolu etylowego. Pułapki były eksponowane w terenie przez 5 miesięcy, od początku maja do końca września, a materiał wybierano z nich w odstępach miesięcznych. Analizę zebranych biegaczowatych przeprowadzono w Zakładzie Zoologii Instytutu Biologii Uniwersytetu Humanistyczno-Przyrodniczego Jana Kochanowskiego w Kielcach.
Do oceny wpływu czynników środowiskowych na faunę biegaczowatych posłużono się odpowiednimi wskaźnikami ekologicznymi: - strukturą dominacji zgrupowania, - wskaźnikiem różnorodności gatunkowej, - wskaźnikiem równomierności, - strukturą troficzną zgrupowania, - wskaźnikiem łowności.
Struktura dominacji zgrupowania. Ustalamy ją, określając dominację indywidualną każdego gatunku za pomocą wzoru
gdzie: n – liczba osobników danego gatunku, N – liczba wszystkich osobników badanej grupy taksonomicznej. W zależności od procentowego udziału danego gatunku w zgrupowaniu, zaliczamy go do określonej klasy dominacji. Przyjęto podział wskaźnika dominacji na 5 klas (Górny, Grüm 1981): D1 – subrecedenty: gatunki, których osobniki stanowią ≤ 1,0% osobników zgrupowania; D2 – recedenty: gatunki, których osobniki stanowią 1,1-2,0% osobników zgrupowania; D3 – subdominanty: gatunki, których osobniki stanowią 2,1-5,0% osobników zgrupowania; D4 – dominanty: gatunki, których osobniki stanowią 5,1-10,0% osobników zgrupowania; D5 – eudominanty: gatunki, których osobniki stanowią > 10,0% osobników zgrupowania.
Różnorodność zgrupowań. Różnorodność zgrupowań oszacowana została przy pomocy wskaźnika różnorodności Shanonna i Weavera (H’)
gdzie
pi oznacza frakcję
Równomierność podziału osobników między gatunki. Oceniona została za pomocą wskaźnika równomierności Shannona-Weavera (J’)
gdzie H’ – wartość wskaźnika różnorodności Shanonna i Weavera, S – liczba gatunków
Analizę zoogeograficzną zebranego materiału dokonano w oparciu o klasyfikację Leśniaka (1987).
4.10.1. Bór bagienny
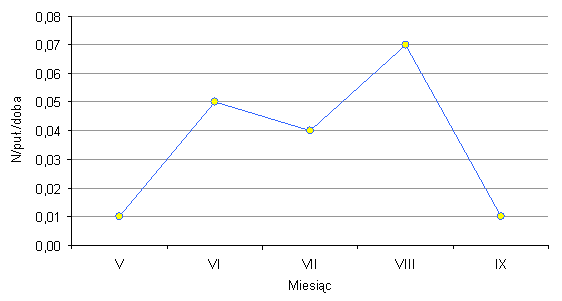
Na powierzchni zlokalizowanej w borze bagiennym odłowiono 85 chrząszczy z rodziny biegaczowatych Carabidae, które należały do 6 gatunków (Tab. 55). W odłowionym materiale zabrakło chrząszczy z gatunku Carabus cancellatus oraz Pterostichus strenuus, które w poprzednim roku był odławiane na tej powierzchni. W zamian pojawiły się dwa inne gatunki: Pterostichus oblongopunctatus oraz P. nigrita (oba jako subdominanty). Średnia łowność biegaczowatych w pułapki Barbera wynosiła 0,04 osobnika na pułapkę w ciągu doby. Przebieg łowności biegaczowatych w ciągu całego okresu badań przedstawia rys. 84. W odłowionym materiale jako eudominaty występowały dwa gatunki - szykoń czarny Pterostichus niger i biegacz fioletowy Carabus violaceus. Szykoń czarny jest leśnym gatunkiem „wszędobylskim” – występuje zarówno w drzewostanach liściastych jak i iglastych. Jego najwyższa aktywność przypadała na lipiec. Biegacz fioletowy, tak jak i pozostałe gatunki z tego rodzaju, objęty jest ochroną ścisłą (Rozporządzenie Ministra Środowiska, 2004). Jest to szeroko rozprzestrzeniony gatunek europejski, najczęściej spotykany w lasach liściastych i mieszanych.
Tabela 55. Charakterystyka gatunków biegaczowatych odłowionych w borze bagiennym w 2009 roku Table 55. Characteristics of catching ground beetles in coniferal swamp forest (2009)
Rys. 84. Przebieg łowności Carabidae w pułapki Barbera na powierzchni w borze bagiennym (Sobolewo 2009) Fig. 84. Course of catching Carabidae in Barber traps on the surface in marshy coniferous forest (Sobolewo 2009)
Struktura dominacji w zgrupowaniu biegaczowatych przedstawia się następująco: - eudominanty (D5) stanowiły 80,0%; - dominanty (D4) stanowiły 7,1% - subdominanty (D3) stanowiły 12,9%.
W strukturze troficznej zdecydowanie dominowały duże zoofagi (Tab. 56), osobniki o ciężarze powyżej 100 mg, odżywiające się głównie pokarmem zwierzęcym. Stanowiły one 91,8% wszystkich odłowionych na powierzchni biegaczowatych. Ponadto występowały małe zoofagi, osobniki odżywiające się również pokarmem zwierzęcym ale o wadze poniżej 100 mg. Stanowiły one 8,2% odłowionych biegaczowatych.
Tabela 56. Struktura troficzna zgrupowania biegaczowatych w borze bagiennym dla dostępnych lat obserwacji Table 56. Trophic structure of the ground beetles’ grouping in marshy coniferous forest for the available observations
Tabela 57. Charakterystyka zgrupowania biegaczowatych boru bagiennego dla dostępnych lat obserwacji Table 57. Characteristics of the ground beetles’ grouping of marshy coniferous forest for the available observations
Zgrupowanie biegaczowatych występujące w borze bagiennym jest dość ubogie – stwierdzono zaledwie 6 gatunków. Wielkość wskaźnika różnorodności Shannona-Weavera wynosi H’=1,792 a równomierności J’=0,100 (Tab. 57).
4.10.2. Las mieszany
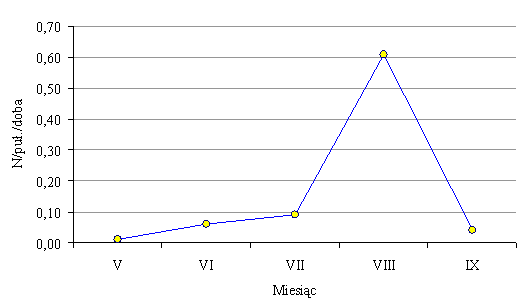
W lesie mieszanym odłowiono łącznie 565 chrząszczy z rodziny Carabidae, należących do 8 gatunków (Tab. 58). W porównaniu do poprzedniego roku w odłowionym materiale nie stwierdzono tylko jednego gatunku - Nebria brevicollis. Pojawiły się natomiast dwa inne: Carabus granulatu i Cychrys caraboides. Średnia łowność biegaczowatych w pułapki Barbera wynosiła 0,25 osobnika na pułapkę w ciągu doby i była ponad sześciokrotnie wyższa, niż w borze bagiennym. Przebieg łowności biegaczowatych w ciągu całego okresu badań przedstawia rys. 85.
Rys. 85. Przebieg łowności Carabidae w pułapki Barbera na powierzchni w lesie mieszanym (Sobolewo 2009) Fig. 85. Course of catching Carabidae in Barber traps on the surface in mixed forest (Sobolewo 2009)
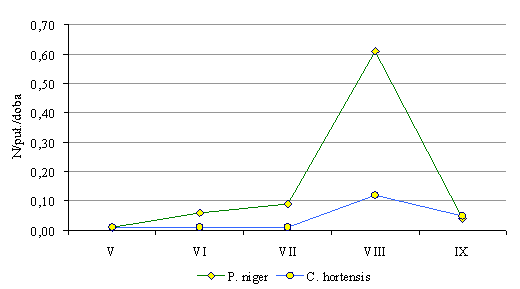
Rolę eudominanta w odłowionym materiale odgrywały dwa gatunki - szykoń czarny Pterostichus niger, tak, jak w poprzednim roku oraz biegacz ogrodowy Carabus hortensis. Szykoń czarny stanowił 66% wszystkich odłowionych biegaczowatych, a jego najwyższa łowność przypadała na sierpień (Rys. 86). Biegacz ogrodowy odławiany był znacznie rzadziej i stanowił ponad 15% wszystkich odłowionych biegaczowatych.
Struktura dominacji w zgrupowaniu przedstawia się następująco: - eudominanty (D5) stanowiły 81,6%; - dominanty (D4) stanowiły 12,6% - subdominant (D3) stanowiły 3,9%, - recendenty (D2) stanowiły 1,4%, - subrecedenty (D1) stanowiły 0,5%.
Tabela 58. Charakterystyka gatunków biegaczowatych odłowionych w lesie mieszanym w 2009 roku Table 58. Characteristics of catching ground beetles in mixed forest (2009)
W strukturze troficznej dominowały, tak, jak i w borze bagiennym, duże zoofagi (Tab. 58, 59). Stanowiły one 93,6% wszystkich odłowionych na powierzchni biegaczowatych. Ponadto występowały małe zoofagi, które stanowiły 6,4% odłowionych biegaczowatych.
Tabela 59. Struktura troficzna zgrupowania biegaczowatych w lesie mieszanym dla dostępnych lat obserwacji Table 59. Trophic structure of ground beetles’ grouping in mixed forest for the available observation years
W lesie mieszanym stwierdzono 8 gatunków biegaczowatych. Wielkość wskaźnika różnorodności Shannona-Weavera wynosi H’=2,079 a równomierności J’=0,313 (Tab. 60).
Rys. 86. Przebieg łowności Pterostichus niger i Carabus hortensis w pułapki Barbera na powierzchni w lesie mieszanym (Sobolewo 2009) Fig. 86. Course of catching Pterostichus niger and Carabus hortensis in Barber traps on the surface in mixed forest (Sobolewo 2009)
Tabela 60. Charakterystyka zgrupowania biegaczowatych lasu mieszanego dla dostępnych lat obserwacji Table 60. Characteristics of the ground beetles’ grouping in mixed forest for the available observations
Literatura
Górny M., Grüm L. 1981. Metody stosowane w zoologii gleby. PWN, Warszawa, s. 483. Leśniak A. 1987. Zoogeographical analysis of the Carabidae (Coleoptera) of Poland. Fragm. Faun., Warszawa, 30, s. 297-312.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||